C4植物の研究者のWebサイトです メールでのお問い合わせはこちら
C4光合成には、3つのパターンあります。
C4植物の行うC4光合成は2種類の光合成細胞,葉肉細胞(MC)と維管束鞘細胞(BSC)から成り立っています.下図はソルガムの断面写真とその模式図ですが、維管束の周りを維管束鞘がとりかこみ、そこに維管束鞘細胞があります。さらにその周りに葉肉組織が存在しています.基本的には下の模式図のような光合成を行っています.C4植物は維管束鞘細胞内の脱炭酸する酵素(下図の赤矢印)の違いによって3つのサブタイプに分かれます。サブタイプそれぞれについてより詳しく知りたい場合はさらにその下をご覧ください.
より詳しい説明
NEDP-ME型
NED-ME型
PEP-CK型
 |
 |
より詳しい説明
気孔から入ってきたCO2は葉肉細胞の細胞質のCarbonic
anhydrase(CO2とHCO3-の平衡反応を触媒する酵素)によってHCO3-となり、次にPEPカルボキシラーゼ(PEPc,酵素No.1)
によってHCO3-がオキザロ酢酸(OAA)として固定されます。ここまでが共通部分です.ここからサブタイプによって異なります.NADP-ME型ではOAAは還元されてリンゴ酸となり、NAD-ME型とPCK型ではアミノ化されてアスパラギン酸(Asp)となります。これらのC4ジカルボン酸はBSCに移動し,脱炭酸酵素によって放出されたCO2はRubiscoによって再固定されてPGAになります。その後C3回路により還元されて糖リン酸となり,ショ糖やデンプンが作らます。さらに脱炭酸された残りのC3化合物は,ピルビン酸リン酸ジキナーゼ(PPDK)によって最初の炭酸受容体であるPEPに再生されます。C4植物はこのようなCO2濃縮機構を持っているため,BSC内のCO2分圧が高く保たれ,Rubiscoのオキシゲナーゼ活性は抑制され,光合成効率は高まっています。CO2分圧が高く保たれるためにはBSCの細胞壁がCO2拡散を抑制する機能を持つことが必要となり,実際,BSCのCO2透過性はC3植物のMCにくらべ約100倍低いことがわかっています(Jenkins
et al.1989)。
ちょっとした疑問:このようにC4光合成の成立には組織分化と機能分化が結びつくことが不可欠です。もし、PEPCとRubiscoが同一細胞内に存在したらどうなるのでしょうか?
- →PEPCの方がRubiscoに比べCO2に対する親和性が高いため,PEPCによって主にCO2が固定され,Rubiscoが働かなくなってしまう.
もし,PEPCと脱炭酸酵素が同一細胞内に存在したらどうなってしまうのでしょうか?→左図のようにATPだけを消費し,CO2が堂々めぐりになってしまうのでしょう.
1990年代からC4光合成に関わる酵素の遺伝子をC3植物に発現させる研究が多数行われています。酵素活性は元のC3植物に比べ,数倍増加していますが,C4植物に比べると著しく低く,光合成能力はほとんど変化しないかあるいは低下しています.C4光合成では組織が分化し,特定の組織に特異的に酵素が発現する必要があるのです。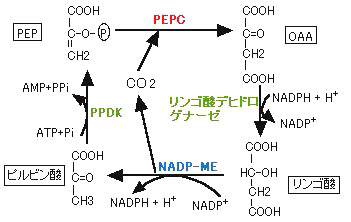
さらに、サブタイプそれぞれについてより詳しく知りたい方はこちら
NEDP-ME型
葉肉細胞(MC)で作られたオキザロ酢酸(OAA)のほとんど(%は種によって異なる)は,これに特異なトランスロケーターにより葉緑体内に輸送され,NADP-リンゴ酸デヒドロゲナーゼ(酵素No.2)によってリンゴ酸(MA)に還元されるが、一部はアスパラギン酸アミノトランスフェラーゼ(酵素No.9)によってアスパラギン酸となる。リンゴ酸(MA)は原形質連絡(Plasmodesmata)を通って維管束鞘細胞に拡散して入り、特定の輸送体により維管束鞘葉緑体に達する.両細胞間でリンゴ酸の拡散流が起こるためには少なくとも約2mMの濃度勾配が必要である.維管束鞘細胞の葉緑体内ではNADP-マリックエンザイム(酵素No.4、このサブタイプの名前の由来)によって脱炭酸され、CO2はRuBP
carboxylase(酵素No.10)の働きによってRuBPに固定され2分子のホスホグリセリン酸となりカルビン回路をへてデンプンなどの炭水化物が合成される。脱炭酸によって生産されたピルビン酸(PA)はおそらく特異的なトランスロケーターにより維管束鞘葉緑体から輸出され,原形質連絡を通ってMCに拡散し、MC葉緑体にある特異的トランスロケーターにより葉緑体内に輸送される.そこでピルビン酸リン酸ジキナーゼ(酵素No.3)によって再びCO2固定の受容体となる。NADP-ME型のある植物ではホスホエノールピルビン酸カルボキシラーゼ(酵素No.6)が2番目の脱炭酸酵素として使われる(Gutierrez
et al.1974, Walker et al.1997)。
NADP-ME型では維管束鞘細胞の葉緑体にグラナがほとんどないため,カルビン回路に必要なNADPHは葉肉細胞の葉緑体の非循環的光合成電子伝達系から提供して補わなくてはならない.その一部はリンゴ酸の酸化的脱炭酸により生じたNADPHを利用するが,残りの部分は両細胞葉緑体の内包膜にあるトリオースリン酸-リン酸トランスロケーターを介したトリオースリン酸-3-ホスホグリセリン酸シャトルにより,葉肉葉緑体から維管束鞘葉緑体にATPとNADPHが共に転送される(ATPは維管束鞘葉緑体の光化学Iの循環的電子伝達光リン酸化によっても供給される).

NED-ME型
葉肉細胞のホスホエノールピルビン酸カルボキシラーゼによって生じたオキザロ酢酸(OAA)は細胞質でグルタミン酸-オキサロ酢酸アミノトランスフェラーゼ(酵素No.9)によってアミノ基が転移されてアスパラギン酸(Asp)に変換される.維管束鞘細胞に拡散したアスパラギン酸(Asp)はトランスロケーターによりミトコンドリアに入り,イソ酵素のアスパラギン酸アミノトランスフェラーゼにより再度、オキサロ酢酸(OAA)となり、NADリンゴ酸デヒドロゲナーゼ(酵素No.7)
によって還元され、リンゴ酸(MA)となり,NAD-マリックエンザイム(酵素No.5、このサブタイプの名前の由来)により酸化的に脱炭酸され、ピルビン酸(PA)になる.このときリンゴ酸デヒドロゲナーゼによってできたNAD+がNADHに再還元される.ミトコンドリアから放出されたCO2は葉緑体に拡散し,RuBP
carboxylase(酵素No.10)によってカルビン回路に取り込まれる。脱炭酸の生産物であるピルビン酸はピルビン酸トランスロケーターを経てミトコンドリアを離れ,細胞質のアラニンアミノトランスフェラーゼ(酵素No.8)によってアラニン(Ala)に変換される.再び葉肉細胞に拡散で戻ったアラニンはアミノトランスフェラーゼのイソ酵素により再びピルビン酸(PA)に変えられ,葉緑体に輸送された後,NADP-ME型と同様にピルビン酸-リン酸ジキナーゼによってホスホエノールピルビン酸(PEP)に変換される。

PEP-CK型
PCK型はBSCの細胞質にあるPEP carboxykinase(酵素No.6)がメインの脱炭酸酵素であるが、BSCのミトコンドリア内にNAD-malic
enzyme(酵素No.5)も持っており、2つ回路で脱炭酸される。MCで asparate aminotransferase(酵素No.9)によって生産されたAspはBSCの細胞質でホスホエノールピルビン酸カルボキシラーゼ(酵素No.6)によって脱炭酸され、CO2はRuBP
carboxylase(酵素No.10)によってCO2カルビン回路に取り込まれる(これがメイン)。MCの葉緑体内でつくられたいくらかのMAはBSCのミトコンドリア内でNAD-malic
enzyme(酵素No.5)よって脱炭酸され、カルビン回路に取込まれる(これがサブ回路であり、NAD-ME型と同じ)。 PCK型植物にはかなりのNAD-ME活性が見出されることがある。単離維管束鞘細胞を用いた実験によりEriochloa
borumensis では脱炭酸反応の70%がPEP-CK、30%がNAD-MEにより行われるという(Rathnam, C. K. M. and
G. E. Edwards 1975)

3つの回路で脱炭酸する場所が違うことに注目される。NAD-ME型とPCK型は葉緑体とミトコンドリアとの共同作業によって光合成を行っていることがNADP−ME型と大きく異なる点である。PCK型では、基質がOAAであることと脱炭酸反応にATPが必要な点が特徴である。
注:番号は酵素名1, PEPcarboxylase; 2, NADP-malate dehydrogenase; 3, pyruvate phosphate dikinase; 3a, adenylatekinase; 3b, pyrophosphatase; 4, NADP-malic enzyme; 5, NAD-malic enzyme; 6, PEP carboxykinase; 7, NAD-malate dehydrogenase; 8, alanine aminotransferase: 9, asparate aminotransferase; 10, RuBP carboxylase; 11, carbonic anhydrase; 12, respiratory electron transport system.