C4�A���̏o���Ƃ��ꂩ��Evolution and Future
C4�A���̏o���@
| �ŏ��̗���A��Cooksonia�̉��́A4���N�ȏ�O�̃E�F�[���Y�̌Â�����猩���������iLang1937�AEdwards��1992�j�AC4�A���̏o���́A����ɔ�ׂ�A���ŋ߂ł���悤���B �@ �@C4�A���͏��Ȃ��Ƃ�30��A18�ȑ��ɔ�є�тɏo�����Ă��邱�Ƃ���A���̌n�������̉ߒ��ő����I�ɐi�������ƍl�����Ă���iMoore1982�j�BC4�A�����ŏ��Ɍ��ꂽ�����ɂ��Ă͏������邪�A���̏؋��Ƃ��āA1250���N�O�̉��iNambudiri1978�j�A�܂��A�E�ʐ^�̂悤�Ȓ��V���i5-700���N�O�j�̃N�����c�\����������������Ă���iThomasson��1986�j�B���Γy��̒Y�f���ʑ̂̕��͂ɂ���Ă������̐A���𐄑����邱�Ƃ��ł��邪�A������̕�������ɔN�オ�����̂ڂ�A1530���N�O��C4�A�������炵���؋��������Ă���iKingston1994�j�BFox��(2003)�́A����ɂ����̂ڂ�A�k�Ă̑呐���n�т̉��Γy�납��A��2300���N�O��C4�A�����������Ղ�F�߂Ă���B |
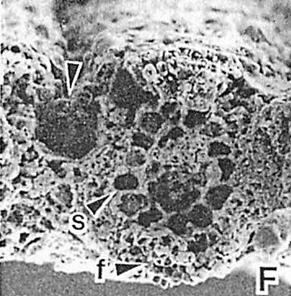 �N�����c�\�����c��t�̉��iThomasson��1986�j |
�@Ehleringer et al.�i1991�j�́AC4�A���̌�������������l���Ĕ����I�ȍ~�i6500���N~�j�ɏo�������ƍl���Ă���B�ߔN�̕��q���v��p������@�ł��A�ŏ���C4�A���̏o���́A5000~6500���N�O�Ɛ��肳��iJacobs��1999�j�A�����Ehleringer��̐����㉟�����Ă���B
�@������6500���N�O�ɐ�ł����Ƃ���ƁA��L�̐��ł́AC4�A���́A���H�����ɐH�ׂ��邱�Ƃ͂Ȃ������ƂȂ��Ă��܂����A�X�E�F�[�f�����R�j�����ق̌����`�[�����A�����C���h��6500-7100���N�O�ɐ����������H���̃e�B�^�m�T�E���X���̋����̕��̉�����C�l�Ȃ̒��Ԃɓ����I�Ȕ����V���J�\�����������Ɣ��\�����iPrasad��2005�j�B�����I�ɂ͂��łɃC�l�ȐA�����o�����A���l���������Ă���A���̑��l���̒���C4�A�����܂܂�Ă���\�����������Ă���BBrown and Smith(1972)���܂��A�C�l�ȐA���̌n����������l�@���čŏ���C4�A���͔����I�ɏo�������ƍl���Ă���B�������Ȃ���A���܂���C4�A�����o�����������ɂ��Ă͌��_�͏o�Ă��炸�A�l�I�ɂ́A�u������C4�A����H�ׂĂ����v�A�uC�S�A���������I����覐̏Փ˂ɂ��u�Փ˂̓~�v���ǂ̂悤�ɏ��z�����̂��v�Ȃǂ��l����Ɗy�����Ȃ�̂ŁAC4�A���A�����I�o�����𐄂������C������B
C4�A���̔����I������CO2�Z�x�@-�r�A�����O�i2007�A�݂������[�j���Q�l��-
| Cerling��(1997)�́A���H�M���ނ̎��̉��͂��āA�����ɂǂ̂悤�ȐA���������Ă������𐄑������i�E�}�j�B�Ȃ��Ȃ玕�̓��ʑ̑g���́A�������H�ׂĂ����A���̓��ʑ̑g���i�{�T�C�g�́y�����z���Q�Ɓj�f���邩�炾�B����ɂ��ƁA800���N�O�܂ŃA�t���J��C���h���嗤�ɂ���V�}�E�}��E�}�̂悤�ȑ��H���M���ނ��H�ׂĂ����̂͂قƂ��C3�A���̎������������A���ꂩ��100���N�̂����ɔނ�̐H�����قƂ�ǂ��ׂ�C4�A���ɕς�������Ƃ������Ă���B �@����܂ł��̂悤�Ȗ�700���N�O�����C4�A���̔����I�����́A��C����CO2�Z�x�̒ቺ�ɑΉ����ċN���������́u��_���Y�f�Q����v�Ǝ����l���Ă����̂����A����ȒP���Șb�ł͂Ȃ��炵���B |
 |
| �����m�쐼���̊C�ꂩ��@�킳�ꂽ�͐ϕ��R�A�̊C�m���A���v�����N�g���iPanani��1999�j�A�C�E�iRoyer��2001�j�A�z�E�f�̓��ʑ́iPearson��2000�j���番�͂��ꂽ��_���Y�f�̋L�^�ɂ��ƁA500���N����1600���N�܂ł̊ԁA��_���Y�f�Z�x�͒n���w�I�ɂ͒Ⴂ�Ƃ����郌�x���������ƕۂ��Ă����炵���i���}�A����C4�A���̔����I�����j�B���Ȃ킿�AC4�A���̑������L�������1000���N�O�����_���Y�f�͒ቺ���Ă����̂��B�ł́A���̎��ԓI�Ȃ���͂ǂ����痈�Ă����̂��H | |
 |
|
| �ŏ��ɐX�����݂��Ă����Ƃ�����A���邢�������D��C4�A���́A�����ȒP�ɈÂ��X�̒��ɓ����Ă����Ȃ��͂��ł���B�ł͂Ȃ��X���Ȃ��Ȃ����̂��A���R�͂킩��Ȃ����A�C���h�����X�[���̉e���������Ȃ��Ċ������A�ȑO�͓~�ɏ\���ȉJ���~�����ꏊ�ʼnJ���~��Ȃ��Ȃ�iQuade��1995�j�A��������c�ꂸ�A�X�т̍X�V���ł��Ȃ��Ȃ����i�r�A�����O2007)�B����ɂ�����ABond��Midgley�i2000�j�́A�u��_���Y�f�Q����v���ǂ��A�����ɂ���Č͂ꂽ�G���|��Ă����Ԃ�C4�A�����N�����A���G�ɂ͔R���₷���Ύ�ƂȂ�A�R�Ύ��������Ȃ������ƂŁAC4�A�����n���s����萶���A�Ă��Ղ߂Đ����A�������g�債���炵���B�m���ɐ��E�̐A���p�^�[�����l�����ʼnΎ��̖����͑傫���AC4�A���̑������p�ɂɉΎ����N�����ĐX�т̔��B��x�点�Ă���Ƃ����؋��̓n���C��j���[�J���h�j�A�Ō������Ă���iHughes��1991�j�B |
�@���̌�́A200���N�ȍ~�̕X���ƊԕX���̎����X����CO2�Z�x�̒ቺ���I�����ƂȂ��ĕX���ɂ����Ĕ�r�I���g�ł������M�ђn����C4�A�������債���ƍl�����Ă���iEhleringer et al.1997�j�B
�n�����g����C4�A��
CO2�Z�x���㏸���鏫��C4�A���͂ǂ��Ȃ�̂ł��傤�ˁBCO2�Z�x�̏㏸�ƂƂ��Ɍ̐��͌�������̂����邢�͉��x�̏㏸�̕����L���ɓ����A�ɉh���Ă����̂ł��傤���H������ƍl���Ă݂܂����B
�@���݁A�l�ނ̊����͂��̕S���N�̊Ԃōł��������x���ł����C��CO2�Z�x������ɑ��������Â��Ă���B��̓I�Ȑ��l�Ō�����1750�`1800�N��CO2�Z�x��280ppm�A���݂ł͖�370ppm�ƂȂ��Ă���B����ɍ����I�̌㔼�ɂ�700ppm�܂ő�������Ɨ\�z����Ă���B
�@�n����̐A�����l���Ă݂�ƁACO2�Z�x�����_�I�ɂ�C4�ւƐi�������I��I��pressure���Ȃ��Ȃ���錻�݂̏ł�C4�A�������ނ��AC3�A���̐A�����L����Ƃ������������邪�A�������ʂɂ��n�\���x��1.5�`4���㏸����ƍl�����Ă���iBolin
et al. 1986, Houghton et al.
1990�j�ACO2�Z�x�����łȂ������̊��v�����ω����A���̗\�z�͑�ύ���ł���B�����ł͍�CO2�����Ő�����C3��C4�A���̔��������������čl���A�����̐A���̐A���ɂ��ė\�����Ă݂����B
1.���ړI�e��
CO2�̔Z�x��350ppm����2�{��700ppm�ɂ����C4�A���̌������͌��݂�CO2�Z�x�ŖO�a���Ă���A�قƂ�Ǐ㏸���Ȃ��BC3�A���̌��������x�͇@Rubisco�̐�����A���ċz���������邽�ߏ㏸����A�܂��A�ACO2���q��Rubisco�̊��������ʂ������邱�Ƃɂ����50�`60���㏸����BC3�A����CO2�Z�x�㏸�𗘗p�ł��邩�ǂ����́A���̏㏸�������������x���Ԏ����ł��邩�ǂ����ɂ������Ă���iKirschbaum
et al.1995,
Bowes1996�j�B�������̈ێ��͂ЂƂ́A�엿�����܂��g���邩�ɂ������Ă���B��CO2�����ŁA�y��ɏ\���̔엿�����������C3�̃o�C�I�}�X�͑�������iPetterson
and Flint 1990, Drake et al. 1996�j���A�s�������ꍇ�ɂ͒����̌������Ɛ����͌�������iBazzaz1990,
Stitt1991, Diaz et al.
1993�j�B����āA������C4�A���͒��f�s���ɋ����̂ŁA��CO2�ɂȂ�A�엿�����������Ȃ����ꍇ�ɂ́AC4�A���̕����L���ɂȂ�̂�������Ȃ��B�܂��A�����ԍ�CO2�Z�x�����ɒu�����ꍇ�A�R���g���[���Ɠ������x������ȉ��̑��x�����߂��ꍇ������i����1988�ACure
and Acock 1986�j�B���̒ቺ�̗v���͇@�f���v���̃N�����v���X�g���~�ςɂ��t�B�[�h�o�b�N�}���iSage et al.1988, Woodrow
and Berry1988�j�A�ARubisco�����̒ቺ�iBaker and Allen 1993, Besford 1993, Rowland-Bamford
et al.1991, Wong 1979, Yelle et
al.1989�j�A�Bsink-source�o�����X�̕���ɂ��V���N���~�b�g�iStitt1991�j���l�����Ă���B
2.�ԐړI�e���P�iCO2�Z�x�㏸���C�E���j
�@CO2�Z�x���㏸�����C3,C4�A���ɂ�����炸�C�E�͕���iAkita
and Moss 1972, Monson1987, Norby and O'Neill 1991, Pallas 1965, Rogers et al.
1983�j�B���̃��J�j�Y���ɂ��Ă͂܂����S�ɉ𖾂���Ă��Ȃ����A���݂̂Ƃ���ACO2���E�ӍזE�̃����S�_�����ɉe�����y�ڂ����A�E�ӍזE�̍זE���ɂ���H+�|���v�ɉe�����y�ڂ����iEdwards
and Bowling
1985�j�A�E�ӍזE��pH�ɒ��ډe�����y�ڂ���(Blatt1987)�Ȃǂ��������Ă���BMonson(1985)�ɂ���CO2�ɑ���C3,C4�A���̔����ɂ͍��͂Ȃ�CO2��2�{�ɂȂ�Ɨ��A���Ƃ���40�����������ƕ��Ă���B�������A�얞��i1996�j�AHollinger�i1987�j�AEamus�i1991�j��̃f�[�^�ɂ��ƋC�E�̔����͎��������ɂ���ėl�X�ɕω�����悤�ł���B
3.�ԐړI�e��2�iCO2�Z�x�㏸���C�E�������U�}�����t�̐��|�e���V�����ێ��j
�C�E�����邱�ƂŁA���U�͗}������A�t�̐��|�e���V�����͍����ێ������iMonson
1993, Rozema et al. 1991, Tyree and Alexander 1993�j�B���̌��ۂ͐��X�g���X�̑ϐ��ɂȂ���iOwensby
et al. 1996, 1997, Chaudhuri et al. 1986, Gifford and Monson 1985, Rogers et
al.1984, Sionit et al. 1981, Tolley and Strain
1984�j�A�������n�тւ̕��z��̊g��Ƌ����͂Ƃ�����������iDahlman 1993�j�B
4.�ԐړI�e��3�iCO2�Z�x�㏸���C�E�������U�}�����t���㏸�j
C3�A���̏ꍇ�A�t���̏㏸�͌��ċz�������邪�AC4�A���ł͉e�������Ȃ��B�܂��A����ňÌċz�����i����邽�߁ACO2�Œ�ɂƂ��Ă̓}�C�i�X�v���ł���B
5.�C���㏸�̉e��
�@�n���̉��g���͍����ɂ��K������C4�A���ɗL���ɂȂ�ƍl�����邪�A���g�̒��x�������Əd�v�Ȃ̂͒g�����Ȃ鎞���ł���B�C��̕ω��̃��f���͓~�G�⍂�ܓx�n��͂�艷�g���̉e�����邱�Ƃ�\�����Ă���iKattenberg
et al.
1996�j�BC3�A���ɂƂ��ē~�⑁�t�͊ɂ₩�ȗ����������ƂȂ�AC3�A���������I�Ȏ����ƂȂ�B�X�^�[�g�_�b�V���ŁA�������Ă��܂��A�n��A�n�����̐��͑����ɏ����Ƃ��ł���B���Ɍ��̋����ɑ��ėL���ƂȂ�ƍl������B
���g���ɂ���āA������傫���ω�����̂��~���ʂł���B�n����̍~���ʂ��邱�Ƃɂɂ���āA�A���������Ă����Ȃ����x���܂ō~���ʂ������Ă��܂��A�����̊ԋC�E���ɂ���ėt�̐��|�e���V�������ێ�����Ă��A�Ӗ��͂Ȃ��B
CO2�Z�x���㏸�����1�`5�̂悤�Ȍ��ۂ��N����B�얞�i1996�j�������������Ƃ���ACO2�{���ɔ����o�C�I�}�X���Y�̕��ϑ���������C3�A����20�`50���AC4�A���ł�9�`14���͈̔͂ɂ������BPooter�i1993�j��156�̐A����Ɋւ��ĕ��������������ʁAC3���41���AC4�A����22���̑����F�߂Ă���B�����̌��ʂ͎�Ƃ��čœK���x�������Ŏ��{���ꂽ���ʂł��邪�AIdso et al.(1987)�͍�CO2�Z�x�̉e���͍�����ԂŊm������邱�Ɩ��炩�ɂ����i19���ȏ�Ńv���X�j�B���̂悤��CO2�Z�x���{������ƍ앨�̎��ʂ�o�C�I�}�X���Y�����傷��Ƃ����������B�������A2050�N�܂łɒn���̐l����50����������ƍl�����Ă���iCassman 1999, Cohen and Federoff 1999, Alexandratos 1999�j�A�앨���Y��CO2�����ɂ�鑝�������ł͂Ƃ��Ă��ǂ����Ȃ����낤�B
C3,C4�A���̐��͑������l���Ă݂�Ɓ|�C�����㏸���ACO2�Z�x�������Ȃ��C3�AC4�A���̌������Ɛ����͏㏸���邪�A���̃M���b�v�͏k�܂邪�A����ł����S�ɂ͂��̍��͂Ȃ��Ȃ�Ȃ��iHunt et al.1996, Grise 1996�j�BC4�A���������ŁA�L���Ȃ܂܂ł��闝�R�̈�͒g�����ꏊ�ɂ��Ή����������̓�����������ł���B
�A���̂Ƃ��Ă͏�L�̂悤�Ȕ����ƂȂ�ƍl�����邪�A�̐��Ƃ���������͈͂ōl���Ă݂��C4�A���͂��̐���͈͂���芣���n��܂ōL���邱�Ƃ��ł���悤�ɂȂ邾�낤�BC3�A�������l�ɂ��̐���͈͂��L���邱�Ƃ��ł���ƍl������B5�ŋ������悤�ɁA���{�̏t�̂悤�ȋG�߂�����n��ł́A�X�^�[�g�_�b�V����C3�A����C4�A�����쒀����n������邩������Ȃ����A�͂����茾���ėv�����������āA�悭�킩��Ȃ��B���_�͂��ꂩ��̌�����҂����Ȃ��B
�Q�l�����FSage
and Pearcy 2000 The Physiological Ecology of C4 Photosynthesis In: Leegood et
al. (eds), Photosynthesis: Physiology and Metabolism pp.497-532 Kluwer Academic
Publishers. ��CO2�Z�x���ƌ����� 1996 �얞�F�M
C4�A���̌����̗��j
C4�A���͈ꕔ�̐A���������ăN�����c�t�\�������A���Ȃ킿�A�t�ǂƓ��ǂ��ʂ�ۊǑ����ۊǑ���זE���Ԋ̂悤�ɕ�͂��A���̊O����t���זE����芪���ėt���̋C����ԂƐڂ��Ă���B���̍\���͂��ł�1884�N��Gustaf haberlandt�������u�����I�A����U�w�v�ɋL�q���Ă��āA�Ƃ�킯�T�g�E�L�r��G���ނ̓��זE�����̃N�����c��z�u���Ƃ�͈̂ۊǑ���זE�Ɨt���זE�̗t�Α̂̊Ԃɂ͉������m�̕��ƓI���������邽�߂ł͂Ȃ����Ƙ_���Ă����B
�@C4�A����C4�A���Ƃ��ĔF�������ȑO�����L�̑��ɂ��AC4�A���̌`�Ԃ̓����Ƃ��Ēm���Ă���ȉ��̂��Ƃ����o����Ă����BRhoades and Carvalho(1944)�̓g�E�����R�V�Ńf���v�����~�ς���ۊǑ���זE�̗t�Α̂ƒ~�ς��Ȃ��t���זE�̗t�Α̂����݂��邱�ƁAHodge�i1955�j�̓g�E�����R�V�̈ۊǑ���זE�̗t�Α̂ɂ͂قƂ�ǃO���i�\�����Ȃ��A�t���זE�̗t�Α̂̓O���i�����B���Ă��邱�Ƃ����Ă����B
�@���łɑS�A���E�ɂ�����J���r���]�x���\����H�̕��ՓI���݂��F�߂����������A�n���C�̃T�g�E�L�r�͔|���������Hugo Kortschak��̓T�g�E�L�r�̗t�ɂ����o�H�������Ă��邱�Ƃ��������ƁA14CO2�������������s�����B�Ƃ��낪�A�ނ�̗\�z�ɔ����āA���̏����Y����PGA�i�Y�f��3�̃��m�J���{���_�j�⒆�ԑ�ӕ��ł��铜�����_�������ł͂Ȃ������B ���̌��ʂ�1954�N�Ɍ������̔N��ɕ��ꂽ�B���̌㐔�N�Ԃ̎����ɂ��T�g�E�L�r�t�ł͌������I14CO2�Œ菉���Y���͂����Ƀ����S�_�̓A�X�p���M���_�Ȃǁi�Y�f��4�̃W�J���{���_�AC�|DCA�j�ł���A�J���r���]�x���\����H�̏����Y���ł���PGA�ⓜ�����_�������́A�ނ���C4����������14C�����A�ŏI�I�ɃX�N���[�X��f���v���������Ƃ������_�ɒB�����B1960�N���V�A��Yuri Karpilov�����l�̂��Ƃ��g�E�����R�V�Ō��o���Ă������A�����̐l�̒m��Ƃ���Ƃ͂Ȃ�Ȃ������B
�@1965�N�A�A�����J�̐A�������w��ɔ��\���ꂽ�ނ�̘_���iKortschak, H. P., C. E. Hartt and G.O. Burr 1965 Plant Physiol. 40: 209- �j�͑傫�Ȕ������ĂыN�������B���N�I�[�X�g�����A�̐�����Ќ�������Hatch��Slack�͂����ǔF���A�����14C�|�Œ�Y������14C���z�ׁA�����n�Ɋ֗^���鏔�y�f�𖾂炩�ɂ���ȂǖȖ��Ȑ����w�I�����������A�����Ɍ�������C4�W�J���{���_�o�H�ƌĂ��V�����Y�_�Œ�o�H������̂ł���B�ނ�͂��̌o�H���������A���̓T�g�E�L�r�̂ق��ɂ��g�E�����R�V�A�J���c���O�T�Ȃǂ̒P�q�t�A���A�A�I�r���Ȃǂ̑o�q�t�A���ɂ����o����邱�Ƃ�������(Hatch, H. D. and C. R. Slack: Biochem. J., 101: 103-)�B�ނ炪���Â���C4�W�J���{���_�o�H�̓n�b�`�E�X���b�N�o�H�Ƃ��Ă��B
C4�������o�H�����ׂĂ�C4�A���ŕK����������łȂ����Ƃ͑�������C�t����Ă����BNADP-ME�^�b4�A���̗t���זE�t�Α̂ɂ�NADP�����S�_�f�q�h���Q�i�[�[�����������A�A�X�p���M���_�A�~�m�g�����X�t�F���[�[�������ア�̂�14CO2�Œ菉���Y���̓����S�_�������Ȃ�BNAD�]ME�A�o�b�j�^�ł͗t���זE�i�זE���j����шۊǑ���זE�ɂ̓A�X�p���M���_��A���j���̃A�~�m�g�����X�t�F���[�[����������14CO2�Œ菉���Y���̓A�X�p���M���_�ł���A3�̃T�u�^�C�v���m�������iHatch et al. 1975�j�܂ł�NADP-ME�^�̓����S�_�^��NAD�]ME�Ƃo�b�j�^�̓A�X�p���M���_�^�ƂȂ��Ă���_�����������B
�@1980�N��ɓ���AHattersley�� �i1983,92�j�͌`�Ԃ��ώ@���钆�ŁAC4�T�u�^�C�v�̌`�Ԃ̈Ⴂ�����o���A���{�̑���3�̃T�u�^�C�v�ɕ��ނ����B�����č��ł͂قƂ�ǂ̑��{��C3�A����C4�T�u�^�C�v�̈�ɕ������Ă� ��iSage et al. 1999�j�B
2001�N�ɂ��`�ԂɊւ��Ă��̂悤�Ș_��������܂����B
�T�^�I��C3�A���ł���^�o�R�Ōs�Ɨt���̖ؕ��Ǝt�����͂�ł���זE��C4�������̓�����������iC4�������ɓ��L�̍y�f�����������j���ƁA�܂������̍זE�Ō������Ɏg����Y�f���q�͋C�E����ł͂Ȃ��ۊǑ��n���狟������Ă��邱�Ƃ��������B��L�̂悤�ȍזE�ł͖ؕ��Ǝt�����瓾����Y�f���q4���܂ޗL�@��������E�Y�_���Č������Ɏg�����Ƃ��ł���B�����āA�l�I�ɉߑ���߂���Ƃ��̌s�A�t���ł̌��ۂ��t�ɍL�������̂�C4�������Ƃ������ƂɂȂ�(���ړI�ɂ��̂悤�Ȑi���̉ߒ��͏ؖ�����Ă��Ȃ�)�B�uCharacteristics
of C4 photosynthesis in stems and petioles of C3 flowering plants�v Juliqn M. H. and W. P. Quick(2001) Nature 415:451-454