C4植物の光呼吸 Photorespiration of C4 plants 2013.11.12
しかし、C4植物でも光呼吸の酵素 1.(赤数字は下記の酵素を示す)RuBP carboxylase/oxygenase (Ogren1984), 3.glycolate oxidase (Servaites et al. 1978), 5.Glycine decarboxylase (Ohnishi and Kanai1983), 6.serine hydroxymethyl transferase (Gardestrom et al.1978),7.hydoroxypyruvate recuctase (Gardestrom et al.1978),9.Catalase (Ohnishi et al.1985)を維管束鞘細胞内にそして8.glycerate kinaseを葉肉細胞の葉緑体内に持っており、Calvin回路が存在する維管束鞘細胞を単離すると、光呼吸活性が認められる(Chollet R. and Ogren W. L.(1975))。つまり、潜在的にはC4植物も光呼吸活性を持っている。したがって、生葉で光呼吸活性が見られないのは維管束鞘細胞内で光呼吸が抑えられているか、あるいは光呼吸によるCO2放出があったとしても葉の外に漏れないうちに葉肉細胞により再固定されているものと思われる。実際、deVeau and Burris(1989)はトウモロコシの成熟葉で18Oの取りこみから光呼吸速度を求め、みかけの光合成速度の11%、5-6週間の植物体で3%であることを報告している。最近ではガス交換速度からも光呼吸の存在が認められている(Marocco et al.1997, 2001)。
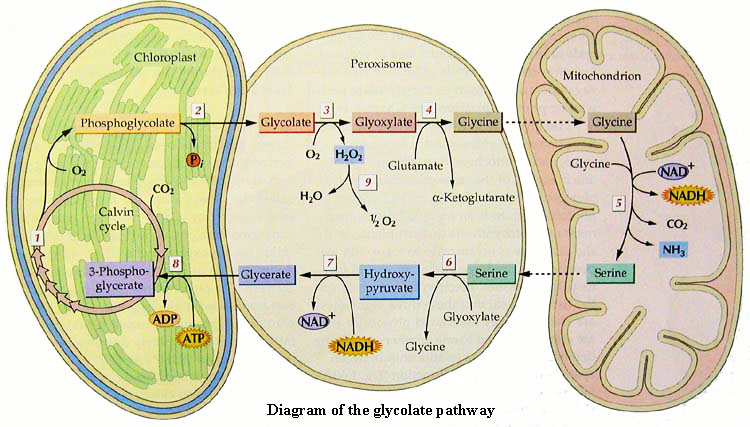
| 1 | Rubulose-1,5-bisphosphate carboxylase/oxygenase |
| 2 | Phosphoglycolate phosphatase |
| 3 | Glycolate oxidase |
| 4 | Glutamate:glyoxylate aminotransferase |
| 5 | Glycine decarboxylase and serine hydroxymethyl transferase |
| 6 | Serine:glyoxylate amino transferase |
| 7 | Hydroxypyruvate reductase |
| 8 | Gycerate kinase |
| 9 | Catalase |
Planta(220:307-317)に掲載された私たちの研究内容ついて書いてみたい。
簡単に言うと形態から見たC4光呼吸活性の評価を行った。光呼吸は上の図のように葉緑体、ペルオキシゾーム、ミトコンドリアの共同作業で行われている。今回は特に、ペルオキシゾームの数、量に注目した。私が考えつくことは当然、先人達も思いつくに決まっており、この種の論文がいくつか発表されている。1971年のFrederick
and Newcombの論文の結果では、細胞の切片当たりのパーオキシゾームの数は、C3植物の方がC4植物より多いこと、C4植物では維管束鞘細胞(BSC)に多く、葉肉細胞(MC)には少ないというもの、すなわち、BSCの方がMCより光呼吸活性が高いことを示している。しかーし、この論文で用いられているC4植物はトウモロコシ(NADP-ME)、スーダングラス(NADP-ME)、ローズグラス(PCK)の3種のみであり、NAD-ME型の植物は測定されておらず、C4植物で一般化するのはちょっと早計と言われてもしかたがない。なぜなら、C4-likeのPanicum
prionitisやEleocharis viviparaの陸上型では、MCのペルオキシゾームの数は、BSCより多く(Brown et al.1983、Ueno
1996)、Frederick and Newcombの結果とは異なっているからである。但し、C3-C4中間種ではC3植物に比べ、MC内のペルオキシゾームの数はBSCより少ない(Brown
et al.1983)。
上で述べたように光呼吸はに葉緑体、ペルオキシゾーム、ミトコンドリアの共同作業で行われいるので、ミトコンドリアの量的な評価をしても面白いのでは?、と考える人もいると思う。確かに植物緑葉ミトコンドリアでは5.Glycine decarboxylase複合体がミトコンドリアのマトリックスに存在し、ミトコンドリア可溶性タンパク質の50%以上を占めている(Walker
and Oliver 1986)ことを考えれば、可能かもしれない。でもミトコンドリアは、呼吸によってエネルギーを取り出す有名な働きもあるので、ペルオキシゾームよりは光呼吸との関わりは少ないような気もする。
ここで、ミトコンドリアの数を評価した論文を見てみると、 Frederick and Newcomb(1971)ではトウモココシ、スーダングラス、ローズグラスで、BSC>MC、Brown et al.(1983)はPanicum属のC3型よりC3-C4中間種ではBSC>MCの差が大きい。しかしC4‐likeのPanicum prionitisではBSCとMCのミトコンドリアの数はほぼ同じである。Ueno (1996)では維管束当たりのミトコンドリアの数はC3型の水生型ではBSC<MCだが、C4‐likeの陸生型ではBSC>MC。Brown
et al.(1989)は、ミトコンドリアとペルオキシゾームの区別がつくような鮮明な電顕写真が撮れなかったのか、この2つの器官の数の合計での評価を行っており、C3型よりC4とC3-C4中間型でBSC内の数が大きいことを示している。このように、ミトコンドリアの数と光呼吸の分布はおよそ対応しているようである。
我々はミトコンドリアの量による評価をさらに進めて、ミトコンドリアの中の5.Glycine decarboxylase(GDC)の量を免疫電顕法によって測定した。その結果、C3植物に比べて量は少ないが、C4植物にも光呼吸が存在することが明らかにされた。また、その活性は3つのサブタイプによって異なり、NADP-ME、PCK、NAD-MEの順に高くなるものと考えられた。
GDC (Glycine decarboxylase)について
GDCはグリコール酸回路の重要な役割を果たす酵素であり、GDCは2モルのグリシンからセリンとアンモニア、CO2を生成する(Oliver,
D. J.
1994)。GDCはかなり大きな酵素の複合体であり、4つの異なったコンポーネント(H、P、T、H-Protein)のたんぱく質からできている。
また、C3-C4中間種やC4植物ではGDCの蓄積は維管束鞘細胞内に限られる(Ohnishi
and Kanai 1983, Hylton et al. 1988, Morgan et al.
1993)。
GDCとともにミトコンドリア内で働くSerine hydroxymethyltransferase (SHMT)
活性もまた葉肉細胞より維管束鞘細胞で多いことがわかっている(Rawsthorne et al. 1988)。
-参考文献の内容を少し-
Ohnishi and Kanai (1983)らは葉肉細胞と維管束鞘細胞に分けてトウモロコシとソルガム(NADP-ME型)、キビ(NAD-ME型)、ローズグラスとギニアグラス(PEP-CK型)の5.酵素Glycine decarboxylaseとSerin hydroxymethltransferaseを測定し、葉肉細胞の活性に比べ、維管束鞘細胞の活性が高いこと、特にNADP-ME型の葉肉細胞における両酵素の活性がほとんどないことを報告しており、葉肉細胞のGlycine decarboxylase活性は、キビ(NAD-ME)、ギニアグラスやローズグラス(PCK)、トウモロコシやソルガム(NADP-ME)の順に光呼吸速度が大きく、サブタイプ間の差異がある可能性を示している。
Usuda and Edwards (1980)は8.の酵素は葉肉の葉緑体にのみ局在していることを証明した。→下記の回路はC3植物の場合のみ成立し、C4光合成ではGlyrateから一度葉肉細胞の葉緑体に入り、それから維管束鞘細胞の葉緑体のカルビン回路に入ることになり、C4植物でのグリコール酸回路はすごく複雑ということになる。
マイクロボディ(ペルオキシゾーム)について
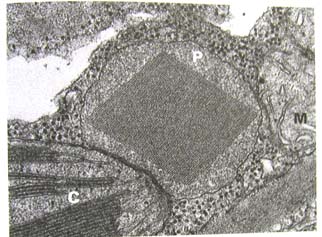 マイクロボディは、真核生物の細胞に存在する細かい顆粒状の基質を一重膜で囲われた直径 0.2〜1.7 μm 程度のほぼ球形のオルガネラである(右写真のP)。C3植物では、発芽時に脂質の流動に関わっており、葉緑体やミトコンドリアと協同して光呼吸に関与し(緑葉ペルオキシソームと呼ばれる-ペルオキシソームと呼ばれる理由は細胞内顆粒。過酸化水素の生成分解に関与する酵素の活性が高いから-)、マメ科の根粒においても重要な役割を果している。植物種や細胞のタイプによっては結晶体や繊維状の内容物を含んでいる。細胞によって数や大きさ、酵素の構成や代謝機能が生育段階や環境条件によって変化する。これは他の真核生物の場合には見られない、植物マイクロボディの特徴となっている。植物と動物ではほぼ50の酵素が局在し、カタラーゼはいつでも存在するので、目印となる酵素となっている。まだまだこのオルガネラの機能については分からないことも多い。(Staehelin
and Newcomb 2000から抜粋)
マイクロボディは、真核生物の細胞に存在する細かい顆粒状の基質を一重膜で囲われた直径 0.2〜1.7 μm 程度のほぼ球形のオルガネラである(右写真のP)。C3植物では、発芽時に脂質の流動に関わっており、葉緑体やミトコンドリアと協同して光呼吸に関与し(緑葉ペルオキシソームと呼ばれる-ペルオキシソームと呼ばれる理由は細胞内顆粒。過酸化水素の生成分解に関与する酵素の活性が高いから-)、マメ科の根粒においても重要な役割を果している。植物種や細胞のタイプによっては結晶体や繊維状の内容物を含んでいる。細胞によって数や大きさ、酵素の構成や代謝機能が生育段階や環境条件によって変化する。これは他の真核生物の場合には見られない、植物マイクロボディの特徴となっている。植物と動物ではほぼ50の酵素が局在し、カタラーゼはいつでも存在するので、目印となる酵素となっている。まだまだこのオルガネラの機能については分からないことも多い。(Staehelin
and Newcomb 2000から抜粋)
ミトコンドリアについて
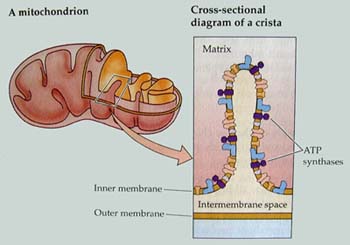 ミトコンドリアはほぼすべての真核生物でみられ呼吸を行なう場所である。ここではTCA回路とそれに伴う電子伝達系ATPの合成反応が行われる.機械的な傷害を受けたり、病原菌に感染したサツマイモ塊根,あるいは吸水発芽時の種子の子葉,胚乳ではミトコンドリアの数の増加が確認されており,活発なエネルギー生産が要求される組織において増大,増加が見られる。直径1μm,長さ1〜3μmであるが、形を変化させたり、原形質流動によって細胞内を活発に移動している.外膜,内膜の2枚の膜を持ち、内膜はひだ状に入り組んでおり、入り組んだ部分をクリステという.内膜の内側には直径6nmほどの粒子が付着しておりその実体はATP合成酵素の一部である。
ミトコンドリアはほぼすべての真核生物でみられ呼吸を行なう場所である。ここではTCA回路とそれに伴う電子伝達系ATPの合成反応が行われる.機械的な傷害を受けたり、病原菌に感染したサツマイモ塊根,あるいは吸水発芽時の種子の子葉,胚乳ではミトコンドリアの数の増加が確認されており,活発なエネルギー生産が要求される組織において増大,増加が見られる。直径1μm,長さ1〜3μmであるが、形を変化させたり、原形質流動によって細胞内を活発に移動している.外膜,内膜の2枚の膜を持ち、内膜はひだ状に入り組んでおり、入り組んだ部分をクリステという.内膜の内側には直径6nmほどの粒子が付着しておりその実体はATP合成酵素の一部である。